In an age of glowing screens and instant digital downloads, the printed book feels almost like an artifact. Yet, it endures—a testament to its tactile pleasure, the scent of its pages, and the timeless ritual of turning them. While digital formats offer convenience, the printed book remains a powerful cultural symbol. For the bookseller, however, this coexistence has created a new set of challenges, transforming their role from mere merchants of books to custodians of a culture, fighting for survival in an increasingly digital world.
The Rise of the Digital Realm: A New Challenger The past two decades have seen a seismic shift in the publishing landscape, driven by two primary forces:
E-books and Digital Readers:
The advent of devices like the Kindle and Kobo made it possible to carry thousands of titles in one’s pocket. For readers who prioritize convenience, storage, and cost, e-books offer a compelling alternative.
Online Retail Giants:
books-online
The sheer scale and speed of online behemoths like Amazon have disrupted traditional retail. They offer a vast catalogue, often at discounted prices, delivered straight to the customer’s doorstep, bypassing the physical bookstore entirely.
This dual assault has left the traditional bookseller on the frontlines of a battle for relevance.
The Bookseller’s Battle: Challenges on Every Shelf The modern bookseller faces a formidable array of challenges that go far beyond just selling books:
Shrinking Margins: The deep discounts offered by online retailers force physical bookstores to compete on price, a losing battle. This erodes their already slim profit margins, making it difficult to cover overheads like rent and staff salaries.
The “Showrooming” Problem: Customers frequently visit bookstores to browse, discover new titles, and get recommendations from staff, only to then purchase the book online for a lower price. The physical store becomes a free showroom for online competitors.
Inventory Management: Deciding what titles to stock is a delicate art. A small bookstore can’t possibly compete with the endless digital catalogue. They must curate a collection that is both commercially viable and reflective of their community’s interests—a constant guessing game.
High Overheads: Brick-and-mortar stores come with significant costs: rent, electricity, maintenance, and staff wages. These fixed costs are a heavy burden, especially with fluctuating foot traffic.
The “Experience” Paradox: While physical bookstores are valued for the browsing experience, community feel, and expert recommendations, translating this intangible value into sales is a constant struggle. The very thing that makes them unique isn’t enough to guarantee profitability.
The Fight for Survival: From Bookstore to Cultural Hub The booksellers who are thriving today have realized that they can’t simply be a place to buy books. They must become something more—a vital cultural hub in their community. Their strategy is a masterclass in adapting to the digital age:
Curated Collections: Rather than trying to stock everything, they specialize. They might focus on local authors, a specific genre (like sci-fi or history), or independent publishers, creating a unique identity that online retailers can’t replicate.
Community Events: Bookstores are becoming venues for author readings, book clubs, poetry slams, children’s story hours, and workshops. These events foster a sense of community and provide a reason for people to step away from their screens and gather.
Expert Recommendations: The biggest advantage a physical bookstore has is its knowledgeable staff. They offer personalized recommendations and a human touch that no algorithm can match.
Partnerships: Collaborating with local cafes, schools, and literary festivals to host events and cross-promote. A bookstore can become a linchpin of the local cultural ecosystem.
Diversification: Many bookstores now sell coffee, stationery, unique gifts, and other merchandise to supplement their income and create a more compelling retail experience.
Conclusion
The printed book is not dead, but it has changed. It is no longer just a vessel for information; it is an object of value, an aesthetic choice, and a symbol of a more mindful way of consuming knowledge. The bookseller is its guardian, a passionate advocate in a world of instant gratification. Their fight for survival is more than a commercial battle; it is a cultural one. By transforming their stores into vibrant community spaces, they are proving that in the digital age, a place where people can gather, browse, and connect over a shared love of reading is more essential than ever. The last page has yet to be turned.
Agriculture has always been the backbone of human civilization. Farmers work tirelessly to grow food for the world, but one of their biggest challenges is protecting crops from harmful pests and insects. Insects such as caterpillars, beetles, aphids, and borers can quickly damage plants, reduce crop yield, and affect overall quality. To tackle this problem, insect killers in agriculture play a vital role. These solutions not only help safeguard crops but also ensure that farmers can achieve better productivity and profitability.
Why Are Insect Killers Important in Farming?
Crops are vulnerable to different pests at various stages of growth. Some insects chew on leaves and stems, while others attack roots or suck plant sap. If not controlled in time, they can cause heavy losses. For example, locusts can destroy acres of farmland within hours, while tiny insects like whiteflies spread plant diseases.
Insect killers provide an effective shield against these threats. By using them properly, farmers can:
Prevent crop damage: Protect leaves, flowers, and fruits from being eaten or destroyed.
Improve yield: Healthy crops produce more harvest, increasing farmers’ income.
Ensure food security: By saving crops, insect killers help in maintaining a stable food supply.
Reduce spread of diseases: Many insects act as carriers of plant diseases, so controlling them reduces overall risk.
type_of_insects
Types of Insect Killers Used in Agriculture
Insect control methods have evolved over time. Today, farmers use a combination of traditional and modern techniques. The major types include:
Chemical Insecticides
These are the most widely used insect killers in farming. They come in different forms such as sprays, powders, or granules. Chemical insecticides act quickly and can control a wide range of pests. However, excessive or careless use can harm the environment, beneficial insects, and even human health. That’s why farmers are encouraged to use them in limited, recommended quantities.
Biological Control
This method involves using natural predators or parasites to kill harmful insects. For example, ladybugs feed on aphids, and certain wasps attack caterpillars. Biological control is eco-friendly and does not leave harmful residues on crops. Many agricultural scientists recommend combining biological control with other methods for sustainable farming.
Organic and Botanical Insect Killers
These are made from natural plant extracts such as neem oil, garlic, or pyrethrum. Organic insect killers are safe for crops, soil, and consumers. They are gaining popularity among farmers who follow organic farming practices. Although they may act slower than chemical insecticides, they are considered healthier and more sustainable in the long run.
Mechanical and Physical Methods
Sometimes farmers use insect traps, sticky sheets, or light-based devices to capture and kill insects. This method is cost-effective and especially useful in greenhouses or small-scale farms.
Integrated Pest Management (IPM)
IPM is a modern approach that combines different methods—chemical, biological, mechanical, and organic—based on the situation. The goal is to reduce dependency on harmful chemicals while keeping pests under control. IPM is widely promoted as a sustainable solution for modern agriculture.
Best Practices for Using Insect Killers
While insect killers are essential, their improper use can cause more harm than good. Here are some best practices:
Identify the pest correctly – Different insects require different control methods.
Use recommended doses – Overuse of chemicals can damage soil fertility and harm beneficial insects.
Follow safety measures – Farmers should wear protective gear when applying insecticides.
Rotate insecticides – Using the same chemical repeatedly can lead to pest resistance.
The Future of Insect Killers in Agriculture
With growing awareness about food safety and environmental health, the future of insect killers lies in eco-friendly innovations. Scientists are developing bio-pesticides, genetically resistant crops, and smart devices that can monitor and control insect populations automatically. These advancements aim to reduce harmful impacts while ensuring strong protection for crops.
Conclusion
Insect killers in agriculture are an essential tool for farmers to protect their hard work and ensure healthy harvests. From traditional chemical sprays to modern organic and biological solutions, these tools provide different options for every farming need. However, the key is to use them responsibly and sustainably. By adopting smart practices and combining multiple methods, farmers can achieve higher yields, protect the environment, and contribute to global food security.
India, a land celebrated for its vibrant culture and diverse culinary heritage, is facing a silent revolution in its kitchens and dining tables. The traditional Indian diet, rich in fresh ingredients, spices, and wholesome grains, is increasingly being sidelined by the allure of readily available, highly processed, and often unhealthy “junk food,” especially among its burgeoning youth population. This shift in food habits carries significant implications for the health and future of the nation.
The Traditional Indian Plate: A Nutritional Powerhouse
Nutritional plate
For generations, the Indian diet has been a testament to balance and nutritional wisdom. Rooted in regional variations, it typically emphasizes:
Whole Grains: Millets like bajra, jowar, and ragi, along with traditional rice varieties and hand-ground wheat, provide complex carbohydrates, fiber, and essential minerals.
Pulses and Legumes: Dals, chana, rajma, and moong are primary sources of plant-based protein, vital for growth and repair.
Fresh Fruits and Vegetables: A kaleidoscope of seasonal produce ensures a rich intake of vitamins, minerals, antioxidants, and dietary fiber.
Healthy Fats (in moderation): Ghee, mustard oil, and groundnut oil, when used appropriately, offer essential fatty acids and fat-soluble vitamins.
Spices: Turmeric, cumin, coriander, and fenugreek aren’t just for flavor; they possess powerful medicinal properties, acting as anti-inflammatories and antioxidants.
Probiotics: Homemade curd and fermented foods support gut health.
This diet, when consumed in traditional portions, naturally provides sustained energy, supports digestion, boosts immunity, and helps in preventing chronic diseases.
The Rise of the “Convenience Culture” and Junk Food
In stark contrast to this wholesome tradition, “junk food” typically refers to highly processed items packed with excessive sugar, salt, unhealthy fats, and empty calories, offering little to no nutritional value. The rapid urbanization, globalization, and aggressive marketing have fueled its meteoric rise in India, particularly among the youth.
Accessibility and Affordability: Fast food outlets are ubiquitous, offering quick, cheap meals that fit into busy schedules.
Taste and Novelty: The high sugar, salt, and fat content in junk food is engineered to be incredibly palatable, creating strong cravings and even addiction.
Peer Influence and Social Status: Consuming branded fast food can be seen as “cool” or a sign of modernity among peer groups.
Aggressive Advertising: Television commercials, social media influencers, and online games constantly promote processed snacks, shaping preferences from a young age.
Lack of Awareness/Time: For busy parents, packaged foods become a convenient option, while some youth may lack adequate knowledge about healthy eating.
The Alarming Consequences for Indian Youth
The shift towards junk food is having a profound and alarming impact on the health of India’s young generation:
Obesity and Overweight: Junk food’s high calorie density and low satiety lead to overeating, contributing to a significant rise in childhood and adolescent obesity. This, in turn, increases the risk of chronic diseases later in life.
Nutritional Deficiencies: Despite consuming more calories, youth consuming junk food often suffer from a lack of essential micronutrients like vitamins, minerals, protein, and fiber, leading to “hidden hunger” and impaired physical and mental development.
Metabolic Disorders: The surge in sugar and unhealthy fat intake is directly linked to an increased risk of Type 2 Diabetes (even in adolescence), high blood pressure, and high cholesterol – conditions once primarily associated with adults.
Digestive Issues: Low fiber content in junk food contributes to constipation, bloating, and other gastrointestinal problems.
Dental Problems: Excessive sugar in soft drinks and processed snacks leads to widespread tooth decay and gum disease.
Mental Health Impact: Emerging research suggests a correlation between a diet high in processed foods and an increased risk of mental health issues like depression, anxiety, and poor concentration. Fluctuations in blood sugar can also affect mood and energy levels.
Reduced Physical Performance: A diet lacking vital nutrients can lead to lethargy, reduced stamina, and overall poorer physical performance in sports and daily activities.
Addressing the Challenge: A Collective Responsibility
Tackling this growing health crisis requires a multi-pronged approach involving government, schools, parents, and the youth themselves:
Government Initiatives: The Food Safety and Standards Authority of India (FSSAI) is at the forefront with initiatives like “Eat Right India.” Campaigns like “Aaj Se Thoda Kam” (promoting reduction in salt, sugar, and fat) and “Trans Fat-Free India” aim to raise awareness. “Eat Right School” integrates nutrition education into curricula, and the “Stop Obesity” campaign (launched in June 2025) targets a nationwide reduction in salt and oil consumption. Regulations on misleading advertising, especially for child-targeted ads, are crucial.
School Policies: Promoting healthier canteen menus, providing nutrition education, and encouraging physical activity within school premises.
Parental Guidance: Parents play a vital role in modeling healthy eating habits, preparing nutritious meals at home, limiting junk food availability, and educating children about mindful eating.
Youth Empowerment: Encouraging critical thinking about food choices, involving them in meal preparation, and highlighting the benefits of traditional Indian foods.
Promoting Traditional Foods: Reviving the consumption of millets, pulses, and seasonal produce through awareness campaigns and making them more accessible and appealing.
The future health of India’s youth hinges on a conscious return to nutritious, balanced diets. While the convenience of junk food is undeniable, understanding its long-term health implications and actively choosing traditional, wholesome options is a battle that needs to be won, one plate at a time.
India, the land of vibrant culture and diverse landscapes, has agriculture as its very backbone. It employs nearly half the workforce and feeds over a billion people. Yet, for decades, Indian farming faced a myriad of challenges: dependence on erratic monsoons, fragmented landholdings, traditional practices, and the looming threat of climate change. Today, however, a quiet revolution is sweeping across the nation’s fields, driven by innovation and technology, promising to significantly increase productivity and secure India’s food future.
The Green Revolution of the 1960s ushered in a new era of high-yield varieties and increased fertilizer use. Now, we are witnessing the dawn of a “Smart Agriculture Revolution,” powered by cutting-edge solutions that are transforming how Indian farmers cultivate their crops, manage resources, and connect with markets.
The Tools of Transformation: How Innovation is Boosting Yields From the smallest plot in Jamshedpur to the vast plains of Punjab, technology is enabling farmers to work smarter, not just harder:
Precision Agriculture: This is the cornerstone of modern agricultural innovation. Using GPS-guided tractors, drones, and IoT sensors, farmers can now get real-time, granular data about their fields.
Soil Sensors: These devices precisely measure soil moisture, nutrient levels, and pH, allowing farmers to apply water and fertilizers exactly where and when needed, minimizing waste and optimizing growth.
Drones: Equipped with multispectral cameras, drones provide aerial insights into crop health, identify pest infestations or disease outbreaks early, and even precisely spray pesticides or nutrients, leading to quicker intervention and reduced crop loss.
Drone
AI-Powered Analytics: Data collected from sensors and drones is analyzed by AI and Machine Learning algorithms to generate actionable insights – predicting yields, advising on optimal planting times, and even creating personalized “crop doctor” apps for farmers’ smartphones.
Polyhouse and Protected Cultivation: Polyhouses (similar to greenhouses) are creating controlled environments for crops. By regulating temperature, humidity, and light, farmers can achieve significantly higher yields, grow off-season crops, and reduce reliance on harmful pesticides. This is a game-changer for enhancing crop quality and extending growing seasons, especially for high-value crops.
Polyhouse
Efficient Water Management (Micro-irrigation): With water scarcity being a critical concern, innovations like drip and sprinkler irrigation are paramount. These systems deliver water directly to the plant roots, drastically reducing water consumption compared to traditional flood irrigation. Government schemes like “Per Drop More Crop” are actively promoting their adoption.
Bio-fortification and Climate-Resilient Crops: Scientists are leveraging biotechnology to develop crop varieties that are more resilient to extreme weather conditions, pests, and diseases. Bio-fortification, which enhances the nutritional value of staple crops, is also crucial for addressing malnutrition.
Farm Mechanization and Automation: Modern machinery, from advanced seeders to mini power weeders, is increasing efficiency and reducing dependence on manual labor. Autonomous machinery and robotic milkers in dairy farming are also emerging, promising to further streamline operations and improve productivity.
Digital Platforms and Market Linkages: Mobile apps and digital platforms are empowering farmers with real-time access to market prices, weather forecasts, expert advice, and government schemes. Platforms like e-NAM are creating online marketplaces, allowing farmers to sell their produce directly to a wider network of buyers, reducing intermediaries and ensuring fairer prices.
Government Initiatives and the Road Ahead The Indian government is actively fostering this agricultural transformation through various initiatives:
Digital Agriculture Mission: Focusing on leveraging technology for transparency and efficiency.
Sub-Mission on Agricultural Mechanization (SMAM): Providing financial assistance for modern machinery and promoting Custom Hiring Centers (CHCs).
Kisan Drones: Subsidies are being provided for the purchase and demonstration of drones in agriculture.
Krishi Vigyan Kendras (KVKs): Providing training, demonstrations, and advisories to farmers at the grassroots level.
Agri-Startups: Nurturing agri-entrepreneurs and supporting innovative solutions.
While challenges remain – including the initial cost of technology, limited digital literacy among some farmers, and ensuring robust rural internet connectivity – the collective efforts of innovators, policymakers, and farmers themselves are steadily overcoming these hurdles.
Conclusion
The future of Indian agriculture is not just about feeding a growing population; it’s about doing so sustainably, efficiently, and profitably for every farmer. By embracing these innovations, India is truly ploughing new ground, transforming its fields into smart, productive, and resilient hubs of growth, ensuring both food security and prosperity for generations to come. The current integration of advanced technologies like AI, IoT, and drones marks a pivotal moment, much like the Green Revolution did decades ago, laying the groundwork for a future where farming is both a science and a sustainable art.
Future Trends in Indian Agriculture Technology
Looking ahead, several exciting trends are poised to further reshape Indian agriculture:
Regenerative Agriculture: An increased focus on practices like no-till farming, cover cropping, and agroforestry to restore soil health, sequester carbon, and enhance biodiversity, moving beyond just productivity to holistic ecological well-being.
Agrivoltaics: The dual use of land for both solar energy generation and agriculture. This can provide farmers with a dual income stream and help reduce water evaporation for crops, especially relevant in sun-rich regions.
Blockchain for Supply Chain Transparency: Deeper integration of blockchain technology to provide immutable records of produce origin, quality, and journey from farm to consumer, building trust and potentially fetching better prices for farmers through direct traceability.
Advanced Biotechnology (CRISPR): Continued breakthroughs in genome editing to create even more resilient, nutrient-dense, and climate-adaptive crop varieties with accelerated breeding cycles, specifically tailored for Indian conditions.
Hyper-local and Controlled Environment Agriculture (CEA): Expansion of vertical farms, hydroponics, and aquaponics, especially in and around urban centers like Jamshedpur, to reduce food miles, ensure fresh produce availability year-round, and minimize land/water usage.
Agri-Fintech Evolution: Greater access to tailored financial products, insurance, and credit for farmers, leveraging AI for risk assessment and personalized offerings, empowering them with better financial stability.
AI-Driven Chatbots and Voice Assistants: More sophisticated AI-powered conversational tools for farmers, providing instant, personalized advice on crop management, pest control, and market access in regional languages, overcoming literacy barriers.
Robotics in Horticulture and Specialized Crops: Development of more specialized robots for delicate tasks like fruit picking, precise pruning, and harvesting high-value crops, where human labor is becoming scarce or expensive.
These trends signify a dynamic and exciting future for Indian agriculture, driven by a continuous cycle of innovation that promises to make farming more intelligent, sustainable, and prosperous.
The role of plants in human health and healing has been unmatched throughout the history of civilization. From the earliest forms of traditional medicine to the most cutting-edge discoveries in modern pharmaceuticals, plants have served as an invaluable source of healing compounds. Among these, phytochemicals — natural chemical compounds produced in plants — have emerged as particularly significant. These compounds possess remarkable anti-inflammatory, antioxidant, and antimicrobial properties. They are demonstrating exceptional potential in the prevention and treatment of a wide range of serious health conditions, including cancer, diabetes, cardiovascular diseases, and liver disorders. Moreover, ongoing research continues to unveil the diverse therapeutic effects of phytochemicals, further cementing their status as vital agents in promoting human health and well-being. As we continue to explore the intricate relationship between plants and health, it becomes increasingly clear that harnessing the power of phytochemicals holds tremendous promise for the future of medicine.
Phytochemicals—including flavonoids, terpenoids, saponins, alkaloids, and many others—exert a wide array of physiological and pharmacological effects on the human body. These naturally occurring compounds, present in plants, have garnered significant scientific interest due to their therapeutic potential. For example, codeine, a well-known analgesic, is derived from the opium poppy (Papaver somniferum) and is extensively utilized for its pain-relieving properties. Similarly, curcumin, the principal active component of turmeric (Curcuma longa), has gained global recognition for its potent anti-inflammatory and anticancer activities. Extensive research on curcumin has demonstrated its efficacy in modulating various cellular pathways, including the reduction of fat accumulation and the enhancement of cellular metabolic processes. These actions contribute to its potential role in the prevention and management of cardiovascular and hepatic diseases. Moreover, curcumin has shown promise in mitigating oxidative stress and inflammation, which are underlying factors in the pathogenesis of numerous chronic diseases. The study of phytochemicals thus continues to open new avenues for developing novel therapeutic agents and nutraceuticals aimed at improving human health and well-being.
The increasing body of research on phytochemicals in the field of medicine has significantly contributed to the development of innovative treatments and therapeutic approaches. Many contemporary pharmacological drugs are now being designed and developed by leveraging the bioactive compounds naturally present in plants, which often exhibit fewer side effects and enhanced efficacy compared to synthetic alternatives. For instance, ginseng—a well-known adaptogenic herb traditionally used to combat fatigue and enhance physical stamina—has demonstrated remarkable potential in both clinical and commercial pharmaceutical applications. Its active components, such as ginsenosides, have been shown to modulate energy metabolism and improve overall vitality, thereby assisting patients in managing fatigue-related conditions. These plant-based medicines offer a more holistic and sustainable approach to healthcare, promoting patient well-being in a safe and long-lasting manner. The integration of phytochemicals into modern drug development not only broadens the scope of treatment options but also reinforces the importance of natural resources in advancing human health.
Botany also plays a pivotal role in indigenous and traditional medicine systems across the world. In India, for instance, the foundations of Ayurvedic and Unani medicine are deeply rooted in the use of plant-based remedies. These traditional medical systems harness the therapeutic properties of various botanicals to address a wide spectrum of health conditions, often without causing significant side effects. For example, Ashwagandha (Withania somnifera), a revered medicinal herb in Ayurveda, is widely recognized for its adaptogenic properties, which help alleviate stress, combat fatigue, and enhance both mental and physical health. This synergy between botany and medicine not only provides effective therapeutic interventions but also plays a crucial role in the prevention of diseases by promoting overall well-being and resilience against stress-related disorders. The continued study and integration of traditional botanical knowledge into modern healthcare frameworks highlight the enduring importance of plants in enhancing human health and quality of life.
Recent research has revealed that certain phytochemicals play a crucial role in protecting human genetic material and preventing DNA damage. This groundbreaking discovery is opening new avenues for the prevention and treatment of cancer, as well as various genetic disorders. For instance, silymarin, a flavonolignan complex derived from milk thistle (Silybum marianum), has shown considerable promise in promoting liver health by protecting hepatocytes from toxins and facilitating tissue regeneration. Such findings underscore the value of integrating botanical and medical research to develop innovative therapeutic approaches. Furthermore, plant-derived compounds have demonstrated efficacy in the treatment and management of a wide array of chronic diseases, including arthritis, diabetes, cardiovascular disease, and more. This highlights the immense potential of natural products in expanding the modern pharmacopoeia, offering safer and often more holistic solutions to complex health challenges. The synergistic collaboration between botanical sciences and modern medicine thus continues to play a vital role in advancing human health and well-being.
With the use of these plant-based medicines, patients not only receive effective treatment but also experience significant improvements in their overall lifestyle and well-being. Consequently, the future of phytochemicals in the medical field is exceptionally promising, capturing the interest of both the general public and scientific communities alike. Modern medical research increasingly confirms that phytochemicals derived from natural plant sources tend to be safer and more beneficial for human health compared to many synthetic drugs and potentially harmful chemicals. This advantage is largely due to their biocompatibility, lower risk of adverse side effects, and their ability to work synergistically with the body’s natural processes. Therefore, further research and development in this field are imperative to establish phytochemicals as a robust and reliable therapeutic option. By deepening our understanding of the mechanisms of action, optimizing extraction and formulation methods, and conducting rigorous clinical trials, the integration of phytochemicals into mainstream healthcare can pave the way for more holistic, effective, and sustainable approaches to disease prevention and management in the future.
It will always be immensely beneficial to understand and utilize this rich treasure trove of plants to support and enhance our health. The integration of medicine and botany not only paves the way for innovative treatment approaches but also significantly advances the prevention of diseases and promotes an overall improvement in quality of life. By harnessing the therapeutic potential of phytochemicals and other plant-derived compounds, we can develop more holistic, safe, and effective healthcare solutions. This collaboration between natural and modern medicine empowers both patients and healthcare providers to embrace a more sustainable and patient-centered approach to wellness. As research continues to unveil the extraordinary potential of plant-based therapies, the synergy of medicine and botany will remain a cornerstone of human health and well-being.
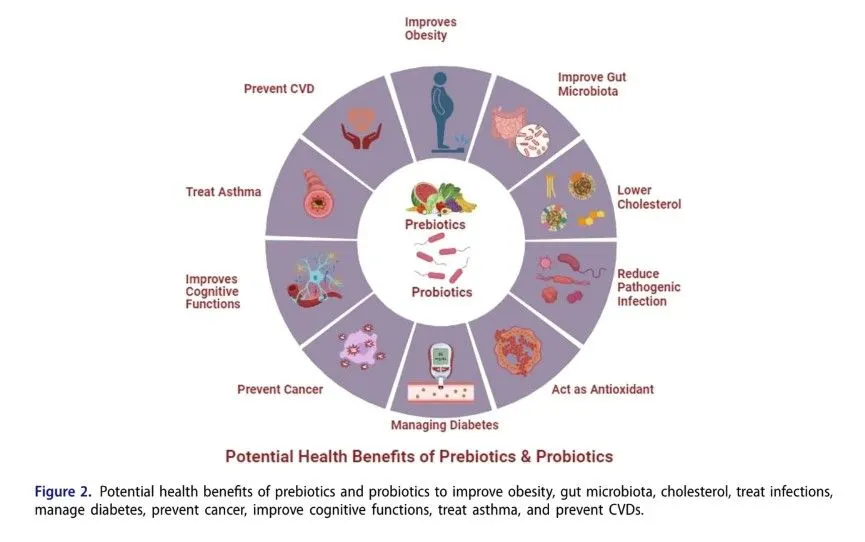
Humans are exceptional reservoirs of diverse microbial species, forming complex microbiota that play a crucial role in health by modulating metabolic processes and protecting against various diseases. The composition and function of the microbiota can be positively influenced by the consumption of probiotics and prebiotics—beneficial bacteria and non-digestible food components, respectively—that promote the growth of these beneficial microbes. Many fermented foods serve as sources of probiotic strains, while plant-based oligosaccharides are well-known prebiotics. Together, probiotics and prebiotics are important in treating immune system disorders, cancer, liver diseases, gastrointestinal issues, type 2 diabetes, and obesity, thanks to their immunomodulatory properties, support of gut barrier integrity, production of antimicrobial compounds, and regulation of immune responses. This review aims to highlight the potential impact of prebiotics and probiotics on gut microbiota, emphasizing their role in enhancing health benefits.
Introduction
The human microbiota contains approximately 101410^{14}1014 bacterial cells, which is comparable to the total number of eukaryotic cells in the human body (Wang et al., 2016). Although the gut of a neonate is initially sterile, it is rapidly colonized by maternal bacteria, resulting in a diverse gut microbiome. During the first few months of life, the infant’s gut microbiota adapts to its environment based on nutritional availability, anaerobic conditions, and microbial interactions (Bäckhed et al., 2015).
The diversity of the adult gut microbiota consists of approximately 1,000–1,150 different bacterial species, primarily including Bacteroidetes, Firmicutes, Actinobacteria, Proteobacteria, and Verrucomicrobia (David et al., 2014; Salvucci, 2019; Almeida et al., 2019). Lifestyle, environment, and age significantly influence the stability of the host microbiome (Di Paola et al., 2011).
Although the gut microbiota remains generally resilient and stable throughout a person’s life, it can be temporarily affected by factors such as unhealthy diets, antibiotic use, and exposure to new environments—though these usually have a limited impact on its overall composition (Rajilić-Stojanović et al., 2013).
As a result, prebiotics are selectively utilized by the host’s microbiota to confer health benefits (Swanson et al., 2020). Their potential effects include modulation of the gut microbiota and the production of beneficial metabolites, such as tryptophan and short-chain fatty acids (SCFAs). Commercially available prebiotics include inulin, isomalto-oligosaccharides (IMO), fructo-oligosaccharides (FOS), galacto-oligosaccharides (GOS), lactulose, and resistant starches (Yan et al., 2018).
Probiotics are live microorganisms that provide health benefits to the host when administered in adequate amounts. Common probiotic species include members of the Bifidobacterium and Lactobacillus genera, while less common probiotics include Faecalibacterium prausnitzii, Akkermansia muciniphila, Streptococcus thermophilus, Saccharomyces boulardii, and Lactococcus lactis (Hill et al., 2014; Markowiak & Śliżewska, 2017; Ballan et al., 2020). These probiotics influence the gut luminal environment, mucosal barrier function, and mucosal immune system.
Composition and diversity of gut microbiota
The human microbiota contains approximately 101410^{14}1014 microorganisms. The mother’s oral microbiota closely resembles that of the placenta, with Firmicutes, Proteobacteria, Bacteroides, Fusobacteria, and Tenericutes contributing to its structure during the prenatal period. According to one theory, microbes may be transferred from the oral cavity to the fetus via the circulatory system (Aagaard et al., 2014).
After birth, the newborn encounters various microbes, with colonization influenced by the mode of delivery (Dominguez-Bello et al., 2010). During vaginal delivery, the baby’s skin and mucous membranes come into contact with the mother’s microbiota, leading to colonization predominantly by Lactobacillus species. In contrast, Propionibacteria and Staphylococcus—common skin microbes—colonize the baby’s mouth, gut, and skin following cesarean section (Jakobsson et al., 2014; Bäckhed et al., 2015).
During the initial months of a newborn’s life, the gut microbiota adapts to its environment, influenced by factors such as nutrient availability, anaerobic conditions, and microbial interactions (Bäckhed et al., 2015). In the first two years, cesarean-born infants tend to have fewer maternally transmitted microbes (e.g., Bifidobacteria and Bacteroides), lower diversity, and a reduced Type 1 T helper (Th1) immune response compared to vaginally born infants; however, these differences gradually diminish over time (Jakobsson et al., 2014; Bäckhed et al., 2015).
Breastfeeding is another key factor in early microbial colonization. Human breast milk contains more than 10710^{7}107 bacterial cells per 800 mL, predominantly from the genera Lactobacillus, Streptococcus, and Staphylococcus (Soto et al., 2014). Additionally, breast milk is rich in oligosaccharides, which selectively promote the growth of Lactobacillus and Bifidobacterium species. These microbes contribute to fermentation in the gastrointestinal tract (GIT), the production of short-chain fatty acids (SCFAs), and the reduction of colonic pH. This acidic environment limits the growth of harmful pathogens that cannot survive under such conditions.
Furthermore, breast milk provides immunoglobulin A, lactoferrin, and defensins, which offer additional health benefits to infants (Lönnerdal, 2016). Early breastfeeding has also been linked to the prevention of diseases such as obesity, dermatitis, and infections (Greer et al., 2008).
Following the introduction of solid foods, the infant’s gut microbiota begins to transition toward an adult-like composition (Turnbaugh et al., 2009). By the third year of life, environmental factors strongly influence microbiota colonization. This increased diversity enhances the synthesis of vitamins and amino acids and improves carbohydrate metabolism (Yatsunenko et al., 2012).
In adulthood, the gut microbiota is typically composed of Firmicutes, Bacteroidetes, Actinobacteria, Verrucomicrobia, and Proteobacteria. This diverse microbiota plays a critical role in human physiology, influencing intestinal barrier integrity, neurotransmitter production, immune system development, and energy metabolism. Lifestyle, physical environment, and age all affect the stability of the host microbiome (Di Paola et al., 2011).
Methodology
This review adheres to PRISMA guidelines and includes literature sourced from Google Scholar, PubMed, Scopus, and Web of Science. Over 50 peer-reviewed studies from the past decade examining the health benefits of probiotics and prebiotics were selected. Quality assessments were conducted using the Cochrane Risk of Bias tool for randomized controlled trials (RCTs) and the Newcastle-Ottawa Scale for observational studies.
The findings were synthesized narratively, focusing on three key areas: the role of gut microbiota in combating diseases, the role of prebiotics in modulating gut microbiota, and the role of probiotics in supporting gut microbiota.
Factors influencing the gut microbiota
To explore the connection between genetics and gut microbiota, researchers profiled the gut microbiota of eight distinct mouse breeds using DNA fingerprinting techniques. A previous study (Kemis et al., 2019) found that the host’s genetic makeup significantly influences microbiota diversity. The host genotype also plays a role in selecting the intestinal microbiota.
At birth, the newborn’s sterile gut is already colonized by numerous microbes from the mother and the surrounding environment. Although the formation of the gut microbiota is influenced by the offspring’s genes, mothers and their children share approximately half of their genetic material as well as similarities in their gut microbiota composition (Coelho et al., 2021).
The adult gut microbiota is highly responsive to dietary changes. In studies where mice were switched to a Western-style diet, the microbiota underwent significant alterations, notably with a marked increase in the abundance of Firmicutes, particularly the class Erysipelotrichi (Salazar et al., 2017). Changes in diet over just 24 hours triggered observable shifts in microbial composition.
Dietary carbohydrates that are indigestible in the upper intestine reach the colon where they are fermented by gut microbes, leading to substantial changes in microbiota composition and beneficial effects on host health (Leeming et al., 2019). The prebiotic hypothesis, first proposed in 1995, highlights that prebiotics can increase the number of Bifidobacteria (phylum Actinobacteria) (Rezende et al., 2021). This microbial shift happens quickly but also reverts rapidly once prebiotic intake stops.
The adult gut microbiota is highly responsive to dietary changes. In studies where mice were switched to a Western-style diet, the microbiota underwent significant alterations, notably with a marked increase in the abundance of Firmicutes, particularly the class Erysipelotrichi (Salazar et al., 2017). Changes in diet over just 24 hours triggered observable shifts in microbial composition.
Dietary carbohydrates that are indigestible in the upper intestine reach the colon where they are fermented by gut microbes, leading to substantial changes in microbiota composition and beneficial effects on host health (Leeming et al., 2019). The prebiotic hypothesis, first proposed in 1995, highlights that prebiotics can increase the number of Bifidobacteria (phylum Actinobacteria) (Rezende et al., 2021). This microbial shift happens quickly but also reverts rapidly once prebiotic intake stops.
Breast milk naturally contains oligosaccharides that act as prebiotics, supporting the growth of Bifidobacterium populations in infants. These findings emphasize the crucial role diet plays in shaping the gut microbiota throughout life (Leeming et al., 2019).
The body’s immune system plays a crucial role in shaping the gut microbiota. Studies have shown that animals with abnormal Toll-like receptor (TLR) signaling exhibit elevated antibody levels, which help regulate commensal bacteria. This interaction between the host and gut microbiota is maintained through increased serum antibody levels.
Mutant mice lacking functional TLRs display altered intestinal microbial compositions, demonstrating that the host’s phenotype is strongly influenced by the characteristics of its gut microbiota. Additionally, factors such as gut peristalsis and the dense mucus layer produced by goblet cells affect the microbial population (Schluter et al., 2020).
The thick mucus layer formed by goblet cells acts as a barrier, limiting microbial penetration into the colonic epithelium (see Figure 1).
Role of microbiota in combating diseases
The gut microbiota plays a key role in metabolic processes, including carbohydrate and lipid metabolism, which are critical factors in the development of diabetes. Certain probiotic strains can improve insulin sensitivity and help regulate blood glucose levels. For example, Lactobacillus and Bifidobacterium strains have been shown to reduce inflammation and enhance glucose metabolism, thereby alleviating the effects of type 2 diabetes (Turnbaugh et al., 2009).
These probiotics exert their beneficial effects by strengthening gut barrier function, lowering endotoxemia, and influencing the secretion of hormones such as incretins, which are involved in insulin release. Moreover, they can affect the gut-brain axis, potentially reducing stress-induced hyperglycemia and further supporting better glycemic control (Schluter et al., 2020).
Microbiota
Gut bacteria metabolize dietary compounds like choline and carnitine into metabolites that influence cardiovascular health. Probiotic strains such as Lactobacillus reuteri help lower cholesterol by breaking down bile acids in the gut, preventing their reabsorption and thereby reducing blood cholesterol levels.
Disruptions in the gut microbiome have been linked to cardiovascular diseases. Certain probiotics offer potential therapeutic benefits by modulating inflammation and reducing hypertension (Leeming et al., 2019). These probiotics also produce short-chain fatty acids (SCFAs), which have been shown to lower blood pressure and enhance endothelial function. Furthermore, they can reduce levels of trimethylamine N-oxide (TMAO), a metabolite associated with higher cardiovascular risk, thus providing a comprehensive approach to supporting heart health (Champagne et al., 2018).
The gut microbiome plays a crucial role in adipose tissue metabolism and the development of obesity. Research shows that the composition of gut microorganisms varies between underweight and overweight individuals. Specific probiotic strains, such as Lactobacillus gasseri and Bifidobacterium breve, influence energy balance and fat storage by regulating nutrient absorption and hormone secretion related to appetite and fat accumulation.
Moreover, short-chain fatty acids (SCFAs) produced by gut bacteria promote adipocyte differentiation, enhance lipid metabolism, and improve insulin sensitivity, all of which contribute to better metabolic health (Aagaard et al., 2014). These SCFAs act as signaling molecules that regulate gene expression involved in lipid metabolism and energy homeostasis. Probiotics also stimulate the release of satiety hormones like peptide YY (PYY) and glucagon-like peptide-1 (GLP-1), which help decrease food intake and support weight loss (Schluter et al., 2020).
Emerging studies indicate that the gut microbiome significantly influences cognitive function and mental health via the gut-brain axis. A healthy gut microbial community is linked to a lower risk of mood disorders such as depression and anxiety. Probiotic strains like Lactobacillus helveticus and Bifidobacterium longum have demonstrated the ability to reduce stress and anxiety symptoms by modulating neurotransmitter production and lowering systemic inflammation (Turnbaugh et al., 2009).
These probiotics impact levels of serotonin and gamma-aminobutyric acid (GABA), both essential neurotransmitters for mood regulation. Additionally, they suppress pro-inflammatory cytokines that can adversely affect brain function. By enhancing gut barrier integrity, these probiotics help prevent inflammatory molecules from reaching the brain, thereby promoting mental well-being (Rosolen et al., 2019).
A diverse gut microbiota is essential for a strong immune system, aiding in the management of asthma and the reduction of allergies. Exposure to a broad range of microbes enhances immune resilience and lowers the risk of autoimmune diseases. Probiotic strains such as Lactobacillus rhamnosus can modulate immune responses and alleviate allergic symptoms by strengthening gut barrier function and decreasing pro-inflammatory cytokines (Aagaard et al., 2014).
These probiotics stimulate the production of regulatory T cells (Tregs), which help sustain immune tolerance and prevent excessive reactions to harmless antigens. They also boost the secretion of secretory IgA, a critical antibody in mucosal immunity, offering extra protection against allergens and pathogens (Schluter et al., 2020).
An imbalance in the gut microbiota is linked to several gastrointestinal disorders, including irritable bowel disease (IBD) and colitis. Managing the gut microbiota through dietary interventions and probiotics can help reduce inflammation and promote gut health. Probiotics such as Saccharomyces boulardii and Lactobacillus plantarum have demonstrated effectiveness in alleviating IBD symptoms and preventing relapses by enhancing mucosal barrier function and modulating immune responses (Leeming et al., 2019).
These probiotics help restore gut flora balance, decrease pro-inflammatory cytokine production, and increase anti-inflammatory cytokine levels. They also support regeneration of the gut epithelium and strengthen gut barrier integrity, thereby reducing intestinal permeability and subsequent inflammation (Champagne et al., 2018).
The gut microbiome plays a crucial role in modulating certain types of cancer, including colon cancer. Some gut bacteria elicit anti-inflammatory responses that may protect against tumor formation. Probiotics such as Lactobacillus casei have been shown to reduce the risk of colon cancer by inhibiting the growth of pathogenic bacteria and promoting the production of anti-carcinogenic compounds (Turnbaugh et al., 2009). These probiotics increase the production of butyrate, a short-chain fatty acid with well-known anti-tumorigenic properties. Butyrate induces apoptosis in cancer cells and inhibits their proliferation. Additionally, probiotics can modulate the immune system to enhance its ability to recognize and destroy cancer cells, providing a dual mechanism against tumor development (Rosolen et al., 2019).
The gut microbiota also influences liver health through several mechanisms. Dysbiosis, or imbalance of gut microbes, can increase intestinal permeability (“leaky gut”), allowing toxins to enter the bloodstream, trigger systemic inflammation, and contribute to liver disorders. Probiotics such as Lactobacillus rhamnosus help maintain gut barrier integrity and reduce liver inflammation. Moreover, gut bacteria participate in bile acid metabolism, essential for digesting dietary fats and regulating lipid and glucose metabolism in the liver (Aagaard et al., 2014). Probiotics can also decrease the production of lipopolysaccharides (LPS), endotoxins that promote liver inflammation and damage. By modulating gut microbiota composition and function, probiotics support liver health and help prevent progression of liver diseases such as non-alcoholic fatty liver disease (NAFLD) and alcoholic liver disease (Rosolen et al., 2019) (see Figure 2).
Role of prebiotics in gut microbiota
Prebiotics are defined as substances selectively utilized by host microorganisms to confer health benefits (Swanson et al., 2020). These benefits include modulation of the gut microbiota and the production of metabolites such as short-chain fatty acids (SCFAs) and tryptophan derivatives. However, these effects should be confirmed in target hosts, including animals and humans (Roager & Licht, 2018; Sanders et al., 2019; Swanson et al., 2020). Common commercially available prebiotics include inulin, lactulose, fructo-oligosaccharides (FOS), isomalto-oligosaccharides (IMO), galactooligosaccharides (GOS), and resistant starch (Yan et al., 2018). Numerous studies have explored the health benefits of dietary fiber consumption, both with and without recognized prebiotic effects. The primary mechanism of prebiotics involves selective fermentation by beneficial gut microorganisms, such as Lactobacillus and Bifidobacterium, which produce acetate and lactate, respectively. These metabolites then stimulate other beneficial microbes to produce butyrate, a key SCFA. Importantly, SCFAs have been shown to enhance mineral absorption, contributing to improved host health (Roager & Licht, 2018; Sanders et al., 2019; Swanson et al., 2020).
Prebiotics can assist in regulating the overall bacterial diversity of the gut by promoting the growth of useful bacteria, while inhibiting the proliferation of potentially dangerous species. Prebiotics can modulate immune responses and reduce inflammation by influencing lymphoid tissue associated with the gut. Prebiotic consumption has been linked to a variety of health benefits, including improved digestive health, enhanced nutrient absorption, and a lower risk of certain chronic diseases such as obesity, diabetes, and cardiovascular disorders.
Health Benefits
According to Oliveira et al., co-cultures of probiotics with certain strains combined with inulin—the most extensively studied prebiotic—improve the acidification rate of dairy products. Santos et al. demonstrated that Lactobacillus acidophilus La-5, when microencapsulated with inulin, showed greater resistance to simulated gastrointestinal tract (GIT) stress in vitro compared to free cells, resulting in an enhanced survival rate (David et al., 2014). Additionally, Rosolen et al. reported that using a combination of whey and inulin as a protective coating for Lactococcus lactis R7 improved heat resistance and tolerance to in vitro GIT stress (see Table 1).
Role of probiotics in gut microbiota
Good health is strongly linked to the ingestion of probiotics. The microbes approved for consumption are generally considered safe, with selective strains targeting specific populations such as newborns, adults, and the elderly. Additionally, the recommended dietary allowance (RDA) of these microbes should be taken into account to achieve optimal health benefits (Hill et al., 2014; Ballan et al., 2020; Coniglio et al., 2023). Common probiotic species include those from the genera Lactobacillus and Bifidobacterium, while other microbes such as Faecalibacterium prausnitzii, Akkermansia muciniphila, Streptococcus thermophilus, Saccharomyces boulardii, and Lactococcus lactis are also categorized as probiotics (Hill et al., 2014; Markowiak & Śliżewska, 2017; Ballan et al., 2020).
Different probiotic strains exhibit varying survival and multiplication rates in the stomach depending on factors such as the food medium (e.g., milk or soymilk), oxygen levels (e.g., stirred yogurt), storage temperature, pH, and the presence of food ingredients or prebiotics (Homayoni Rad et al., 2016; Champagne et al., 2018; Ballan et al., 2020). By reducing certain unfavorable food components, such as raffinose and stachyose found in soymilk, probiotics can exert beneficial health effects (Albuquerque et al., 2017; Battistini et al., 2018; Champagne et al., 2018). While starter strains like Streptococcus thermophilus are added alongside probiotic cultures to shorten fermentation times, their presence can inhibit the production of flavors generated by acetic acid when co-cultured with Bifidobacterium strains (Tripathi & Giri, 2014; Oliveira et al., 2009; Champagne et al., 2018).
Prebiotics are often consumed together with probiotics to form synbiotics, which can reduce fermentation time and enhance the survival rate of probiotics throughout the gut (Oliveira et al., 2009; Markowiak & Śliżewska, 2017). Dietary changes modulate the behavior of probiotic strains differently. For example, the addition of fruit pulp to soymilk fermented with probiotics significantly influences the properties of the final product (Peters et al., 2019). Furthermore, advancing food technologies such as microencapsulation have greatly improved fermentation methods and increased tolerance to gastrointestinal tract (GIT) stresses (Oliveira et al., 2009; Champagne et al., 2018; Tripathi & Giri, 2014). Recent studies indicate that microbial metabolites contribute to health benefits and influence probiotic function and supplementation strategies (Champagne et al., 2018; Kalita et al., 2023; Mehmood et al., 2023) (Table 2).
Mechanism of action
Probiotic microorganisms influence the host in several ways, enhancing the intestinal lumen, mucosal barrier, and immune stability (Fong et al., 2020). These effects are mediated through various cell types involved in both innate and adaptive immunity, including epithelial cells, monocytes, dendritic cells, B cells, T cells (such as regulatory T cells), and natural killer (NK) cells. The primary mechanisms include selective utilization of prebiotics by commensal microbiota, production of metabolites like short-chain fatty acids (SCFAs) and organic acids, reduction of lumen pH, increased mineral absorption, and inhibition of pathogenic growth (Peters et al., 2019) (Figure 3).
Probiotics enhance phagocytosis, regulate immunoglobulin production, improve immune responses, and maintain microbiome homeostasis through competition for nutrients and adhesion sites, bacteriocin release, reduction of pro-inflammatory activities, and enhancement of barrier functions (Bermudez-Brito et al., 2012). Key regulatory pathways and cytokines involved include G protein-coupled receptors (GPR41 and GPR43), glucagon-like peptide 1 (GLP-1), peptide YY (PYY), lipopolysaccharides (LPS), nuclear factor kappa B (NF-κB), tumor necrosis factor-alpha (TNF-α), exopolysaccharides (EPS), interferon-gamma (IFN-γ), and interleukin-12 (IL-12). These mechanisms play important roles in reducing metabolic endotoxemia and inflammation (Peters et al., 2019).
Moreover, probiotics modulate mucosal cell interactions and maintain cellular stability by improving intestinal barrier function. They achieve this by regulating the phosphorylation of cytoskeletal and junctional proteins, which supports barrier integrity through processes such as mucus production.chloride and water secretion, and tight junction protein interactions (Yadav & Jha, 2019).
Enhanced mucosal barrier function is crucial in managing disorders such as inflammatory bowel disease (IBD), celiac disease, gut infections, and type 1 diabetes (Ghosh et al., 2021). At the molecular level, epithelial cells respond differently to commensal or probiotic bacteria compared to pathogens. For instance, probiotic bacteria do not induce interleukin-8 (IL-8) secretion from epithelial cells, whereas pathogens like Shigella dysenteriae, enteropathogenic Escherichia coli, Listeria monocytogenes, and Salmonella dublin do (Bermudez-Brito et al., 2012). In fact, co-culture with….
Probiotic bacteria can reduce IL-8 release caused by these pathogens, thereby mitigating inflammation and promoting intestinal homeostasis. However, not all probiotics exhibit this anti-inflammatory trait; for example, Escherichia coli Nissle 1917 has been shown to increase IL-8 secretion in a dose-dependent manner, highlighting the variability in the immunomodulatory effects of different probiotic strains (Wen et al., 2020).
Future perspectives
Advancements in gut microbiome profiling tech-niques will enable personalized approaches to gut health interventions. By identifying an individual’s gut microbiota composition and its response to
probiotics_machanis
By leveraging prebiotics and probiotics, healthcare professionals can design targeted treatment strategies to maximize health benefits. Researchers continue to explore novel prebiotic and probiotic strains to optimize their effects on gut health. Advances in microbial engineering and genetic editing technologies have facilitated the development of more precise and potent prebiotics and probiotics, thereby maximizing their therapeutic potential (Wen et al., 2020).
The gut-brain axis—a bidirectional communication network linking the gut and the brain—illustrates how gut microorganisms influence mental health and cognition. This connection opens avenues for developing prebiotic and probiotic interventions aimed at supporting mental health and reducing symptoms of depression. Moreover, the therapeutic applications of prebiotics and probiotics extend well beyond gut health. Emerging research has examined their roles in managing metabolic disorders, cardiovascular diseases, and autoimmune conditions. Identifying specific microbial strains and bioactive compounds capable of modulating disease-related pathways offers promising new directions for targeted therapies (Peters et al., 2019).
Microbiome-based therapeutics, including fecal microbiota transplantation (FMT) and defined microbial cocktails, show considerable promise for treating gastrointestinal disorders and systemic diseases. As our understanding of the distinct functions of various microbial communities deepens, these therapies are expected to become increasingly refined and widely accepted in mainstream medicine. However, the rapid growth of prebiotic and probiotic products in the market has outpaced regulatory oversight, leading to concerns about inconsistent quality and efficacy. Enhanced regulatory frameworks will be essential to ensure the safety, reliability, and therapeutic value of these products.
Notably, early-life exposure to prebiotics and probiotics may have long-lasting effects on gut health and overall well-being. Understanding how these early interventions shape the developing gut microbiome—and consequently influence lifelong health—is a critical area of ongoing research (Rosolen et al., 2019).
As gut microbiota science advances, it is likely that nutritional guidelines will increasingly incorporate prebiotic and probiotic recommendations to promote gut health. Integrating these into standard dietary advice could help prevent gut-related disorders and improve general health outcomes. In conclusion, the future of prebiotics and probiotics in gut microbiota research is promising and holds vast potential to enhance human health. Continued exploration of the gut microbiome’s complexities and its broad impact on well-being will drive the development of personalized interventions and innovative therapeutics, revolutionizing approaches to gut health and disease management (Wen et al., 2020).
Limitations
The potential impact of probiotics and prebiotics in enhancing health benefits is promising, yet several limitations must be acknowledged. Much of the current evidence relies on studies with small sample sizes, short durations, or investigations focused on specific populations, thereby limiting the generalizability of findings. Additionally, the variability in probiotic strains, prebiotic compounds, dosages, and methodological inconsistencies across studies complicates the interpretation and comparison of results. Individual differences in microbiome composition and the complex interactions between probiotics, prebiotics, and host physiology are often oversimplified, further challenging the establishment of universal guidelines. Potential biases, such as industry sponsorship and publication bias, can skew outcomes. Moreover, the long-term effects and safety profiles of these interventions are not well-documented, and significant translational gaps remain between research evidence and practical clinical recommendations.
Conclusion
The gut microbiome, a complex network of microorganisms, plays a crucial role in various physiological processes, including nutrient metabolism and immune system regulation. Prebiotics are indigestible food components that stimulate the growth and activity of beneficial microbes in the gut. Through fermentation, prebiotics produce short-chain fatty acids (SCFAs), which confer anti-inflammatory properties and support gut barrier integrity. By promoting the proliferation of beneficial microbes, prebiotics contribute to a healthier gut ecosystem and may protect against gastrointestinal disorders. When consumed in sufficient quantities, probiotics—live beneficial bacteria—can enhance gut barrier function, produce antimicrobial compounds, and modulate immune responses. Probiotics have been shown to alleviate intestinal disorders such as irritable bowel syndrome and antibiotic-associated diarrhea, as well as improve immune function and reduce the risk of infections. Thus, both prebiotics and probiotics play significant roles in improving quality of life by enhancing overall health.
The present study is an attempt to establish a fast, highly reproducible transformation with a simplifed regeneration system in soybean targeting the apical meristem. The modifed half-seed explants from soybean cultivar (cv.) JS335 were subjected to diferent time intervals of sonication (0, 1, 10, 20, and 30 min) and vacuum infltration (0, 1, 10, 20, and 30 min) in the presence of Agrobacterium tumefaciens strain EHA105 harbouring pCAMBIA1301. The explants were then co-cultivated and subjected to a modifed plant regeneration process that involves only two steps (1) primary shoot regeneration, and (2) in vitro rooting of primary shoot. The rooted plantlets were hardened and maintained in the greenhouse until maturity. Sonication treatment of 10 min, followed by plant regeneration using a modifed method, recorded the highest transformation efciency of 26.3% compared to other time duration tested. Furthermore, 10 min of vacuum infltration alone resulted in even higher transformation efciency after regeneration, reaching 28.0%. Interestingly, coupling sonication and vacuum infltration for 10 min respectively produced the highest transformation efciency after regeneration of 38.0%. The putative transformants showed gus expression in mature leaves, trifoliate leaves, fowers, and pods. The presence of hpt II was also confrmed in putative transformants, with an amplicon size of 500 bp. Quantitative real-time PCR confrmed the existence of hpt II as one to two copies in the soybean genome of T0 plants. Furthermore, the segregation pattern was observed in the T1 generation soybean plants which were confrmed using PCR for hpt II. The optimized protocol when tested with other Indian soybean cultivars showed an enhanced transformation efciency ranging from 19.3% (cv. MAUS47) to 36.5% (cv. CO1). This optimized protocol could provide a reliable platform to overcome the challenges that are associated with the genetic engineering of soybean.
Introduction
Soybean (Glycine max (L.) Merrill), an economically valuable crop, is largely used for consumption and industrial applications (Widholm et al. 2010). The global population growth and the consistent demand for soy products are leading to a continuous increase in the production and demand for soybeans. Consequently, signifcant eforts have been dedicated to improving the regeneration system and the efectiveness of transforming soybeans. These eforts show great potential for developing superior soybean varieties with desired characteristics. Up until now, soybean regeneration has been achieved through somatic embryogenesis, direct organogenesis, and indirect organogenesis. However, poor regeneration has been a major obstacle in the indirect organogenesis method for soybeans. Most of the research conducted on soybeans has focused on somatic embryogenesis or direct organogenesis. Regarding soybean transformation, various intrinsic factors such as Agrobacterium strains, types of explants, composition of culture media, duration of co-cultivation, and plant selection markers have been extensively investigated to enhance the efciency of the transformation process. Moreover, extrinsic factors like physical wounding of explants, sonication, and vacuum infiltration have been optimized to achieve higher transformation efciency in soybeans Despite various studies aimed at improving soybean transformation efciency using Agrobacterium infection, the success rate has been very low due to genotype dependency and low regeneration of transformants (Kumari et al. 2016; Liu et al. 2004). Moreover, poor shoot elongation and long regeneration duration are other important limiting factors for the efective regeneration of transformants (Ma and Wu 2008). Thus, there is an urgent need to look for alternative ways to develop transformed soybean to meet the global demand. In this regard, the current study aims to develop a fast, reliable, and efcient soybean transformation system incorporating sonication and vacuum infiltration thereby targeting the apical meristem of modifed half-seed explants. Moreover, the highlight of the present study is the hassle-free and fast regeneration of transformed plants from infected half-seed explants using a simplifed regeneration method that involves just two steps (1) primary shoot regeneration, and (2) in vitro rooting of primary shoot. The optimized protocol has also been tested with 10 cultivars to check its efficiency.
Materials and methods
Indian soybean cultivars (cv.) JS335, PUSA 9712, CO1, TAMS-38, JS71-05, JS93-05, NRC7, MAUS47, PK416, and Punjab 1 were procured from ICAR-Indian Institute of Soybean Research, Indore, Madhya Pradesh, India, and the cultivars were grown and maintained at the research garden, Department of Biotechnology, Bharathiar University, Coimbatore, Tamil Nadu, India. The optimization was carried out using the soybean cultivar (cv.) JS335 (Fig. 1a). To begin the experiment, the seeds of soybean cv. JS335 were subjected to surface sterilization and imbibed in sterile water for a period of 24 h (Fig. 1b). Following imbibition, the seed coat was removed, and the cotyledons were separated. Only the cotyledon containing the embryonic axis was utilized for the study. Additionally, the radicle of the embryo, which was attached to the cotyledon, was carefully dissected to obtain the modifed half-seed explant (Fig. 1c). For primary shoot regeneration, explants were inoculated on MS medium supplemented with diferent concentrations of 6-Benzylaminopurine (BAP) (0–8.8 μM) and cultured for 30 days. The explants were sub-cultured into a fresh medium with respective hormonal concentrations at 15 days intervals. For rooting of primary shoots, MS medium supplemented with diferent concentrations of Indole-3 butyric acid (IBA) (0–9.8 μM) was used and the culture was incubated for 30 days. In-order to select the primary shoot after transformation, minimum inhibitory concentration (MIC) was determined in modifed half-seed explants by inoculating in regeneration medium (MS+2.2 μM BAP; pH 5.7) with diferent concentrations of hygromycin B (0–5 mg l −1) and incubating for 30 days. In addition, the explants were sub-cultured at 15 days intervals. Subsequently, the established primary shoots were transferred to a rooting medium (MS+4.9 μM IBA; pH 5.7) with diferent concentrations of hygromycin B (0 to 3 mg l −1) and incubated for 30 days for selection at the rooting stage. All the cultures were maintained at 25±2 °C under a 16/8-h photoperiod.
Agrobacterium tumefaciens strain EHA105 harbouring pCAMBIA1301 was used for transformation (Fig. 2). The T-DNA region of the binary vector contains hygromycin phosphotransferase II (hpt II) as the plant selection marker and gus as a reporter gene. The vector backbone carries the neomycin phosphotransferase II (npt II) for bacterial selection. Agrobacterium culture was prepared by inoculating a single colony into 30 ml LB broth containing antibiotics such as kanamycin (50 mg l −1) and rifampicin (25 mg l −1). The culture was incubated at 28 °C for 16 h at 180 rpm. The bacterial culture was centrifuged at 6000 rpm for 15 min, and the pellet was suspended in a liquid MS medium. Additionally, a flter-sterilized solution of 200 μM acetosyringone was added to the bacterial suspension, which was then incubated for 1 h at 28 °C at 180 rpm. The absorbance of bacterial suspension was adjusted to 1.0 at OD600 prior to infection. For genetic transformation, the modified half-seed explants were inoculated into 30 ml Agrobacterium suspension and sonicated for diferent durations (0, 1, 10, 20, and 30 min). Similarly, the explants were subjected to vacuum infltration for diferent time intervals (0, 1, 10, 20, and 30 min) in 30 ml Agrobacterium suspension. Finally, the explants were subjected to combined treatments of sonication (10 min) and vacuum infltration (10 min) in the presence of Agrobacterium. After diferent treatments, the explants were then incubated in fresh Agrobacterium suspension (30 ml) at 28 °C for 30 min. After 30 min, the explants were blot-dried and placed in a co-cultivation medium (MS + 200 μM acetosyringone; pH 5.7) and cultured in complete darkness at 25±2 °C for 3 days. Subsequently, the explants were then thoroughly washed with sterile distilled water containing 350 mg l −1 cefotaxime and cultured in regeneration medium (MS + 2.2 μM BAP + 3 mg l−1 hygromycin B; pH 5.7) for 30 days for selection of primary shoots. The excised primary shoots were then cultured ina rooting medium (MS medium+4.9 μM IBA +2 mg l−1 hygromycin B; pH 5.7) for 30 days. The rooted plantlets were carefully removed from the medium, washed with sterile distilled water, hardened for 2 weeks in paper cups and
soybeansoyabean diagram
Mean values of three independent experiments (±) with standard errors (n=100×3). Values with the diferent letters within columns are signifcantly diferent according to Duncan’s multiple range test (DMRT) at a 5% level
aTotal number of primary shoots survived on regeneration medium (MS+2.2 μM BAP+3 mgl−1 hygromycin B) after 30 days of culture.
bTotal number of primary shoots responded for the root development after 30 days of culture on rooting medium (MS+4.9 μM IBA+2 mg l−1 hygromycin B)
cTotal number of putatively transformed plants that survived in the greenhouse after hardening
dTotal number of putatively transformed plants showing the presence of hpt II
eTransformation efciency=number of hpt II PCR positive plants/total number of infected explants×100
The superscript letter f, g, h, i, j shows that these values are signifcantly diferent according to DMRT
twice to ensure accuracy and reliability. As for the transformation experiments, 100 explants were employed for the respective treatments, and the experiments were repeated three times. The resulting data were presented as mean values with the standard error (SE). Statistical analysis was performed using SPSS software version 20, specifcally employing Duncan’s multiple range test (DMRT) to determine signifcant diferences at a signifcance level of P<0.05. For segregation ratio analysis, the SE and Chisquare analysis were used (Gomez and Gomez 1984; Hada et al.2018). Signifcance was determined for values with a P<0.05.
Results and discussion
We have conducted studies on various parameters to improve transformation after regeneration efciency in Indian soybean cultivars to address the challenges associated with soybean transformation, including low regeneration rates and the absence of cultivar-independent protocols. In the soybean direct organogenesis system, the explants will be initially subjected to multiple shoot induction. Then attempts will be made to elongate the shoots, and after elongation, the shoots will be cultured for in vitro rooting. In addition, this process takes approximately 3 or more months to obtain an in vitro rooted plantlet that will be ready for hardening. In order to achieve regeneration using a direct organogenesis system, the radicle, and plumule of the half-seed explants have to be excised and need to be placed in cytokinin containing medium to trigger the meristematic cells to produce multiple shoots. However, in this method, most of the shoots will fail to elongate affecting the regeneration response (Ether et al. 2013). To overcome this limitation with half-seed explants, we have modified the explant preparation in a way that we removed only the radicle and left the plumule intact to obtain the modifed half-seed explants. The presence of plumule in the modifed half-seed explants directed the regeneration system towards primary shoot development followed by subsequent in vitro rooting diverting it from the conventional direct organogenesis process that includes multiple shoot induction, shoot elongation, and rooting. Moreover, this diversion also bypassed the shoot elongation step which was critical in afecting the regeneration efciency. In the present investigation, the primary shoots developed and elongated in the same BAP medium avoiding the necessity of a separate shoot elongation process. Also, using this method, we were able to produce rooted plantlets that are ready for hardening within 60 days (30 days for primary shoot regeneration and 30 days for rooting) which was comparatively less than similar reports on soybean. The aforesaid advantages of using
leave
leave details
Fig. 4 GUS analysis and molecular confrmation of putative transformants regenerated from modifed half-seed explants infected with Agrobacterium tumefaciens strain EHA105 harboring pCAMBIA1301. a Expression of gus in mature leaf from putatively transformed plants; b mature leaf from non-transformed plant; c expression of gus in trifoliate leaves from putatively transformed plants; d trifoliate leaves from non-transformed plant; e expression of gus in fower from putatively transformed plants; f fower from non-transformed plant; g expression of gus in pod from putatively transformed plants; h pod from non-transformed plant; i molecular confirmation for the presence of hpt II in putatively transformed soybean plants. Lane 1: DNA ladder (1 Kb); lane 2: pCAMBIA1301 plasmid (positive control); lane 3: soybean genomic DNA from non-transformed plants (negative control); lanes 4–8: genomic DNA from putatively transformed soybean plants with expected amplicon (500 bp) of hpt II
this modifed half-seed method also highly favoured efcient regeneration in transformation experiments
In the present study, modifed half-seed explants produced the highest response in inducing the primary shoot regeneration (93.0%) in MS medium supplemented with 2.2 µM BAP (Supplementary Table 1). In addition, the maximum in vitro rooting of primary shoots (75.5%) was observed in the MS medium supplemented with 4.9 µM IBA (Supplementary Table 2). Our fndings were similar to those of Arun et al. (2015) and Chen et al. (2018), where the aforementioned concentrations of BAP and IBA showed the best response in inducing shoots and roots in soybean. The MIC of hygromycin B in primary shoot regeneration was found to be 3 mg l−1 and the MIC of hygromycin B in in vitro rooting of primary shoots was 2 mg l−1. The use of hygromycin B as a potent plant selection marker in soybean was established by Olhoft et al. (2006).
In the present study, modifed half-seed explants produced the highest response in inducing the primary shoot regeneration (93.0%) in MS medium supplemented with 2.2 µM BAP (Supplementary Table 1). In addition, the maximum in vitro rooting of primary shoots (75.5%) was observed in the MS medium supplemented with 4.9 µM IBA (Supplementary Table 2). Our fndings were similar to those of Arun et al. (2015) and Chen et al. (2018), where the aforementioned concentrations of BAP and IBA showed the best response in inducing shoots and roots in soybean. The MIC of hygromycin B in primary shoot regeneration was found to be 3 mg l−1 and the MIC of hygromycin B in in vitro rooting of primary shoots was 2 mg l−1. The use of hygromycin B as a potent plant selection marker in soybean was established by Olhoft et al. (2006).
In this study, the transformed modifed half-seed explants were successfully regenerated using an optimized regeneration method. Among the diferent treatments applied, the modifed half-seed explants that underwent a 10-min sonication treatment exhibited the highest number of primary shoots that survived (54.6), along with a substantial number of rooted shoots (44.3) and plants that survived after a 2-week hardening period (26.3). The transformation efciency for this treatment was calculated to be 26.3% (Table 1). However, it is worth noting that the transformation efciency decreased when the sonication time was reduced to 1 min or increased to 20 and 30 min, as indicated in Table 1. The highly active and rapidly dividing meristematic cells that were used for genetic transformation are present in the primary shoot. Sonication creates microwounds through which Agrobacterium could easily reach the meristematic cells and enhance the transformation efficiency (Trick and Finer 1997). Our study is consistent with the fndings of Hada et al. (2018), where the optimum sonication time was found to be 10 min, and increasing the sonication time decreased the transformation efciency from 36.2% to 12.1%. Guo et al. (2015) also claimed that sonication for 2 s improved transformation efciency in soybean from 2.5 to 5.7%. Vacuum infltration has been well validated as an efcient method to improve the rate of transformation by creating negative atmospheric pressure, enabling easy passage for Agrobacterium to target the meristematic cells (Subramanyam et al. 2013). Among diferent time durations for vacuum infltration tested (0, 1, 10, 20, and 30 min), a treatment duration of 10 min showed the maximum number of survived primary shoots (51.6), number of rooted shoots (40.6), number of plants that survived after 2 weeks of hardening (28.0), with the transformation efciency of 28.0% (Table 1). Furthermore, it was observed that extending the vacuum infltration time beyond 10 min had a negative impact on the transformation efciency. This decrease in efciency can be attributed to the injury caused to the explants due to excessive vacuum. Similarly, reducing the vacuum infltration time to 1 min resulted in a decreased transformation efciency of 11.6%. In the case of the treatment involving sonication for 10 min combined with vacuum infltration for 10 min, the number of primary shoots that survived in the selection medium was recorded as 54.6 (Fig. 1d and e). Additionally, the number of rooted shoots was 46.6 (Fig. 1f and g). Following the hardening process, a total of 38.0 plants successfully survived and acclimatized (Fig. 1h and i). Importantly, the transformation efficiency significantly improved to 38.0%, which is considerably higher than the efciency observed in the explants treated with sonication or vacuum infltration alone (Table 1). These fndings align with the previous studies conducted on soybeans by Arun et al. (2015) and Hada et al. (2018), which also suggested that combining sonication and vacuum infltration can enhance the transformation efciency, as demonstrated in the present study.
From the histochemical GUS assay, it was observed that the mature leaf (Fig. 4a), trifoliate leaves (Fig. 4c), flower (Fig. 4e), and pod (Fig. 4g) from putative transformants developed an intense blue colour and tested positive for gus expression. The mature leaf (Fig. 4b), trifoliate leaves (Fig. 4d), fower (Fig. 4f), and pod (Fig. 4h) from nontransformed plants did not show the gus expression. In the present study, the transformation efciency was calculated based on the presence of the hpt II in transformed plants. The T0 plants that survived after hardening were subjected to this analysis. The amplicon size of 500 bp (Fig. 4i, Lane 4–8) indicated the presence of hpt II in transformed plants. pCAMBIA1301 plasmid served as the positive control (Fig. 4i, Lane 2) whereas non- transformed plants did not show any amplifcation for hpt II (Fig. 4i, Lane 3). Overall maximum transformation efciency of 38.0% was achieve when modifed half-seed explants were subjected to sonication (10 min) and vacuum infltration (10 min). In this present study, the copy number of hpt II in T0 plants was determined by quantitative real-time PCR using
table
Mean values of three independent experiments (±) with standard errors (n=100×3). Values with the diferent letters within columns are signifcantly diferent according to Duncan’s multiple range test (DMRT) at a 5% level
aTotal number of primary shoots survived on regeneration medium (MS+2.2 μM BAP+3 mg l
−1 hygromycin B) after 30 days of culture
b Total number of primary shoots responded for the root development after 30 days of culture on rooting medium (MS+4.9 μM IBA+2 mg l−1 hygromycin B)
cTotal number of putatively transformed plants that survived in the greenhouse after hardening
dTotal number of putatively transformed plants showing the presence of hpt II
eTransformation efciency=number of hpt II PCR positive plants/total number of infected explants×100
Actin as the reference gene. The results revealed that the copy number of hpt II ranged between one and two. The T0 transgenic soybean lines GmJS335-2 and GmJS335-3 had two copies, while lines GmJS335-1, GmJS335- 4, GmJS335-5, GmJS335-6, GmJS335-7, GmJS335-8, and GmJS335-9 had one copy of hpt II (Supplementary Table 3). The quantitative real-time PCR is replacing the traditional method of detecting the copy number of the foreign gene via the southern blot technique due to various advantages such as accuracy, lower cost, higher stability, and ease of operation. This technique has been successfully employed in several crops, including cotton (Yang 2012), wheat (Gadaleta et al. 2011), rice (Wei et al. 2011), maize (Yuan et al. 2010), tomato (Wang et al. 2011), and soybean (You-wen et al. 2012). Additionally, the segregation pattern observed in the T1 generation demonstrated Mendelian inheritance with a ratio of 3:1 in one plant (GmJS335 8) at a signifcance level of 0.05% (Supplementary Table 4). The optimized protocol has been further applied to evaluate the transformation efciency in other cultivars of soybean. In the present investigation, cv. JS335 was found to be having the highest transformation efciency of 38.0% followed by the cv. CO1 (36.5%) and cv. TAMS-38 (33.6%). Among the diferent cultivars examined, cv. MAUS47 displayed the lowest transformation frequency, recorded at 19.3% (Table 2). Overall, the method developed in this study proved to be fast and highly efcient in obtaining transgenic lines within a relatively short duration of 60 days to obtain rooted plantlets. In comparison, previous studies conducted by Arun et al. (2015), Hada et al. (2018), and Wang et al. (2022) demonstrated soybean transformation systems utilizing conventional direct organogenesis, which required longer regeneration times of 123 days, 104.5 days, and 97 days, respectively. Therefore, this simplifed transformation and modifed regeneration protocol can be utilized effectively for developing transgenic soybean varieties with desired traits.
Conclusion
In this study, we have successfully developed a simple regeneration system from modifed half-seed explants, consisting of two steps: primary shoot regeneration and rooting. This system has been efectively adapted to regenerate transgenic plants from modifed half-seed explants infected with Agrobacterium, and it ofers the advantage of a shorter regeneration period. Additionally, the incorporation of sonication and vacuum infltration techniques has signifcantly enhanced the transformation efciency in our study. Moreover, this transformation and regeneration system has demonstrated its efcacy across various soybean cultivars, indicating its wide applicability. We believe that this simple protocol holds great potential for commercial trait improvement in diverse soybean varieties.
Supplementary Information The online version contains supplementary material available at https://doi.org/10.1007/s13205-023-03715-8
Acknowledgements The authors are thankful to the University Grants Commission: Basic Startup Research Grant (No.F.30-410/2018 (BSR); dt.: 29.06.2018), New Delhi, Government of India, for fnancial assistance provided to Muthukrishnan Arun, and also thanks to ICAR -Indian Institute of soybean research, Indore, Madhya Pradesh, India for providing soybean seeds to carry out the research work.
Author contributions MA and KS: Conception and idea. KS: performed the experimental work with the assistance of MA. NV and KS wrote the manuscript with the assistance of MA. KS and NV: preparation of tables and fgures. CA: helped in performing experimental analysis. PG and CA: helped in the critical reviewing of the manuscript. Finally, all the authors approved for the manuscript.
Data availability Data generated in this work is included in the manuscript and in the supplementary material. This will be made available on request.
Declaration
Conflict of interest There are no conficts of interest to declare.
Ethical approval for involving human participants and/or animals Not applicable, since this article does not contain any studies with human participants or animals performed by any of the authors.
The agricultural sector is undergoing a technological transformation with the rise of smart farming. Also known as precision agriculture, this method leverages modern technologies like the Internet of Things (IoT), sensors, and drones to optimize farming operations and increase crop productivity.
Smart farming uses real-time data to monitor soil moisture, crop health, and weather patterns. With this information, farmers can make more informed decisions, such as the ideal time for irrigation, fertilization, and harvesting. This precision not only boosts yields but also reduces water usage and waste.
Agricultural drones play a crucial role by capturing aerial images and mapping farmland to detect areas with pests, disease, or nutrient deficiencies. This targeted approach allows for the efficient application of resources, leading to healthier crops and reduced input costs.
IoT devices connected to smartphones give farmers remote access to field conditions, enabling them to manage their farms more effectively. This is particularly beneficial for large-scale farms or areas with limited labor availability.
In addition to increased efficiency, modern agriculture technology helps tackle climate change by minimizing the environmental impact of farming activities. Reduced chemical usage and optimized resource allocation lead to a lower carbon footprint.
In summary, smart farming is revolutionizing the way we cultivate crops. By embracing technology, farmers can enhance productivity, sustainability, and profitability, making precision agriculture the future of farming.
In recent years, organic farming has gained significant attention as a vital part of sustainable agriculture. This eco-friendly approach eliminates the use of synthetic fertilizers, pesticides, and genetically modified organisms (GMOs), focusing instead on natural processes and biodiversity to maintain soil health and productivity.
One of the biggest benefits of organic farming is the improvement of soil health. Through techniques like crop rotation, composting, and green manure, organic farmers enrich the soil without chemicals, making it more fertile and resilient over time. This not only increases yields in the long run but also ensures better water retention and resistance to drought.
Another major advantage is the positive impact on human health. Organic produce is free from harmful chemical residues, making it safer for consumers. Moreover, it often contains higher levels of antioxidants and essential nutrients compared to conventionally grown food.
From an environmental standpoint, eco-friendly farming reduces pollution, conserves water, and enhances biodiversity. It supports a natural balance of insects and organisms, which helps control pests without chemical intervention.
Despite these benefits, organic farming requires more labor and planning, which can make it slightly costlier. However, the long-term gains in soil fertility, environmental health, and product quality outweigh the initial investment.
In conclusion, organic farming plays a crucial role in building a healthier, more sustainable future. As awareness grows, both farmers and consumers are embracing this green revolution, paving the way for a more eco-friendly agriculture system.